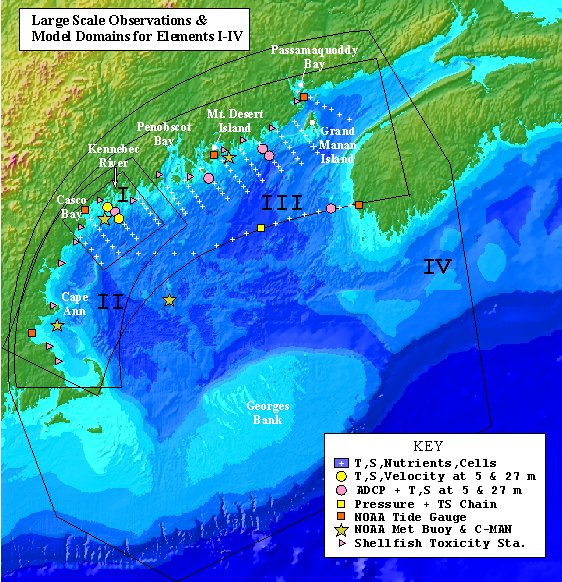
C.3.5.1 Modeling the WMCC. Modeling is a key component of ECOHAB, and ECOHAB-GOM is fortunate to have a robust model on which to base its program. In a study by several of the PIs, a Blumberg and Mellor (1987) 3-dimensional circulation model was developed for the WMCC and Massachusetts Bay (Model Element II, Fig. 5). The objectives of that modeling effort were: 1) to simulate the transport and water properties of the WMCC during 1993-94 as functions of surface, boundary and topographic forcing; and 2) to test hypotheses concerning the mechanisms for initiation, growth, accumulation and transport of Alexandrium cells.
Simulations show many of the observed elements of the WMCC behavior, including trapping along the coast during downwelling and offshore spreading during upwelling. It is evident that wind forcing is an important and possibly dominant source of variability in the WMCC structure. A simple model for Alexandrium growth in response to temperature and salinity was then coupled to the physical transport model. The 1993-94 field seasons were then numerically hindcast using realistic bathymetry, measured wind, river and heat flux data, and boundary conditions derived from our in situ observations. One hypothesis tested was that differences in wind forcing and river discharge caused the large differences in toxicity that were observed in the WMCC between 1993 and 1994. In both years, however, the simulations show low-salinity lenses containing Alexandrium cells traveling along the coast and into Mass Bay, whereas this only happened in 1993. (Movies of model output: http://crusty.er.usgs.gov/ecohab/). Toxicity in the WMCC thus cannot be attributed solely to physical variability of wind and river discharge - it is also heavily dependent on biological variability.
One cause of biological variability is in the process by which Alexandrium cysts inoculate overlying waters. Several hypotheses were thus tested, including the location of the germination input (all along the coast versus discrete seedbeds), but only the scenario in which Alexandrium cells were seeded near the Kennebec river mouth generated patterns of cells that resembled field data. The simulations provide yet another independent indication that Casco Bay is a source region.
In the simulations, cells were continously introduced into the source region at fixed concentrations, giving realistic results for some situations, but not for others. The source function was clearly over-simplified and the modeling team concluded that without a realistic, time-varying input (based on estimates of the germination rates and the duration of germination events), further progress was unlikely in modeling PSP in the WMCC region. Therefore, ECOHAB-GOM will parameterize cyst dynamics in the Casco Bay initiation zone and will incorporate rate data into our existing model. The WMCC model will be expanded to include Casco Bay and the EMCC region, as the latter might be a late-season source for Alexandrium cells. The possibility that established populations from the Bay of Fundy are seeding the EMCC will also be examined.
{kind=link}