C.3.5 Alexandrium and the WMCC. Within the region from the western edge of Penobscot Bay to Massachusetts Bay (Fig. 1), a coupling has been inferred between the distribution of Alexandrium and a buoyant coastal current that travels from east to west. A conceptual model of this linkage has been proposed, termed the "plume advection hypothesis" (Franks and Anderson 1992a). Critical features of this model include: 1) a source population of cells located near the Androscoggin-Kennebec estuary; 2) freshwater outflow from that estuary, resulting in a coastally-trapped, buoyant plume or coastal current (the WMCC) which supports the growth of Alexandrium cells and transports them to the south and west; and 3) plume behavior as influenced by the volume of freshwater outflow, local wind stress, and underlying GOM circulation, all of which combine to regulate the along- and cross-shore location of the plume, its associated cells, and PSP.
The dynamics of this plume and its Alexandrium cells were investigated in a series of small-scale surveys (Franks and Anderson 1992a), and in two major field investigations covering the entire WMCC region and Massachusetts Bay (Geyer et al. submitted ms., Anderson, unpub.). It is now clear that Alexandrium cells are most abundant within the plume, and that downwelling-favorable winds accelerate the plume toward the west and trap it against the shore, resulting in a rapid progression of PSP from east to west. Upwelling winds move the plume offshore, spreading it into a thin, broad layer (Franks and Anderson 1992a; Keafer and Anderson 1993). Persistent upwelling has typically been associated with a decrease in PSP and often in termination of the bloom.
The Alexandrium cells that populate this coastal current presumably originate from germinated cysts, but the location of cyst "seedbeds" and the mechanisms underlying germination remain unknown. Several older cyst surveys document a widespread distribution within nearshore and offshore areas of the GOM, although the offshore survey went no further north than Portsmouth NH (Lewis et al, 1979; Thayer et al, 1983; Anderson and Keafer 1985). Highest concentrations were in the deeper basins where germination appears to be regulated by an endogenous annual clock that restricts excystment to a seasonal window (Anderson and Keafer, 1987). It is not known whether deep water cysts are a major factor in initiating Alexandrium blooms in either the EMCC or the WMCC. Although they are abundant, offshore cysts will germinate gradually in the cold and dark, whereas less numerous cysts in shallow waters may provide a more significant inoculum if their germination is synchronized and rapid, as in Cape Cod salt ponds (Anderson and Keafer, 1985). Offshore cyst deposits might thus be sinks where cysts accumulate through time and germinate too slowly to have a major impact on nearshore blooms. One ECOHAB-GOM objective is to map cyst distributions and identify sites of active excystment which are important in bloom initiation. The rates and duration of cyst germination will also be determined to characterize this important feature of bloom initiation.
It is now clear that there is a source region or "initiation zone" where motile cells of Alexandrium accumulate at the start of the WMCC bloom season. The existence of a major initiation zone in the Casco Bay area is supported by several independent observations. First, seeding of the WMCC with Alexandrium populations from the EMCC is unlikely because the timing is backwards relative to the direction of water flow - i.e. there is a mis-match between the early-season toxicity in the WMCC and the late-season toxicity of the EMCC given that water flows from the latter towards the former (Figs. 1,2). Second, the WMCC is bounded on its upstream side by the toxin-free center of the "PSP sandwich", as well as by the zone where the EMCC deflects from shore. These features represent a barrier delineating the end of one biogeographic region and the beginning of another. Third, our 1993-94 field studies both document the general absence of Alexandrium cells at all stations except those in or immediately downstream from Casco Bay in mid-April (Fig. 4A). The evidence is clear that the Casco Bay region is critical in Alexandrium dynamics in the WMCC, yet the mechanisms underlying this role are not known. A major project focus will be to determine how this region serves as a major source zone for the Alexandrium blooms that populate the WMCC.
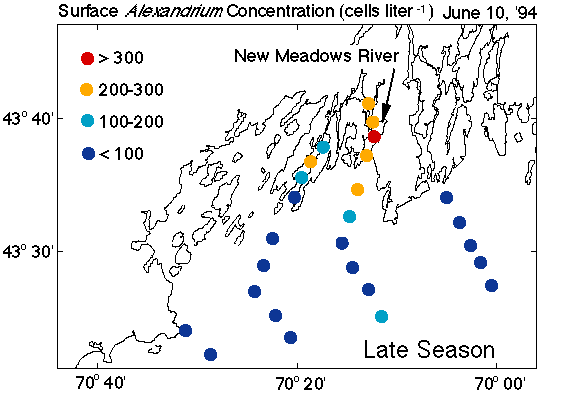
The role of Casco Bay in bloom initiation is only half of the story, however. The persistent westerly flow of the WMCC raises a significant question: If Alexandrium cells are transported to the west each year with no return flow, how does the system "restart" itself from cysts each subsequent spring? With no re-supply of cells to this source region, the PSP problem "downstream" should gradually diminish and disappear, yet PSP in the WMCC has been recurrent nearly every year since 1972 and shows no signs of abatement. A re-seeding mechanism is necessary. Field surveys in 1993 and '94 documented late-season populations of Alexandrium, (such as that in the New Meadows River of Casco Bay in June; Fig. 4C), long after an earlier pulse of cells had been carried to the west by the WMCC. With reduced rainfall and no snowmelt to drive the coastal current and flush the source region, transport was limited and cells could remain in the Bay without washout. Casco Bay thus appears to be the location where late-season, localized blooms persist to re-seed the system with cysts. The mechanisms that regulate these two opposing processes (early season delivery and late season retention) are of fundamental importance and thus will be subject of intense investigation in ECOHAB-GOM.
Summarizing, evidence is strong that the WMCC Alexandrium populations originate near the mouth of the Kennebec estuary, with little or no input of cells from eastern Maine, at least in the early spring. Offshore cyst seedbeds exist, but their quantitative importance remains uncertain. A shallow water "source region" near the origin of the coastal current has been identified, but mechanisms for cell accumulation there remain unknown, as do the conditions that transport water and cells into the WMCC. The origin of the cysts that germinate to produce the inoculum cells is unknown, as are the rate and duration of germination. We believe that cells from the source region are entrained into the coastal current in an episodic, pulsed fashion in the spring, but those mechanisms are unknown. Later in the summer, blooms are retained in that same Casco Bay area such that cyst deposition provides the cells needed to initiate blooms in subsequent years. Behavioral and hydrographic mechanisms that regulate this retention and localization are not known. Overall, the western GOM Alexandrium dynamics have been well-studied and a conceptual understanding of the system exists. Considerable work remains, however, to resolve the many unknowns listed above.
{kind=link}