C.5.2.3 Cyst Dynamics (PIs: Anderson, Keller).
Rationale: A key feature of Alexandrium blooms is their reliance on a resting stage or cyst. Winter temperatures in the GOM are too low to support populations of vegetative cells, so blooms are entirely dependent on seasonal re-inoculation of the water column through the germination of cysts. Our ability to understand and model bloom initiation is dependent upon the distribution of cysts and the rates and duration of the excystment process. Likewise, the timing and magnitude of cyst formation can have a major impact on bloom decline. Since bloom initiation and decline are key features of ECOHAB , this project will examine cyst distribution and dynamics in considerable detail.
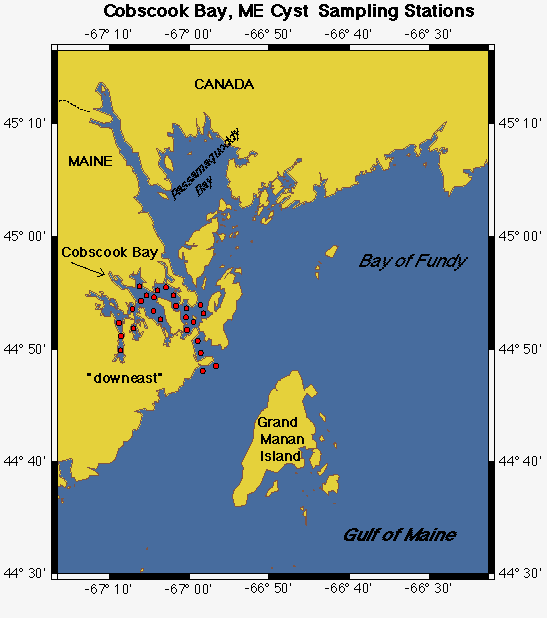
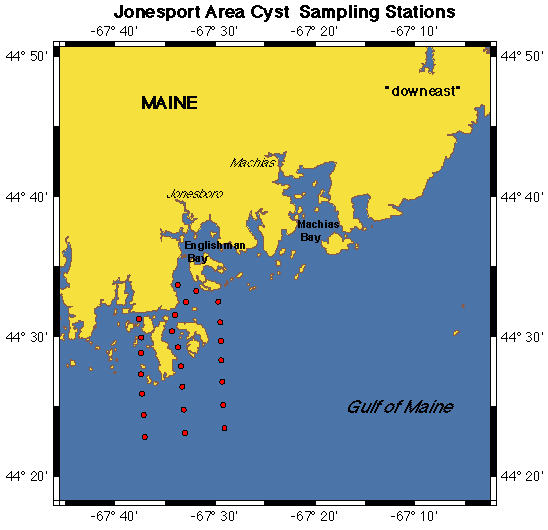
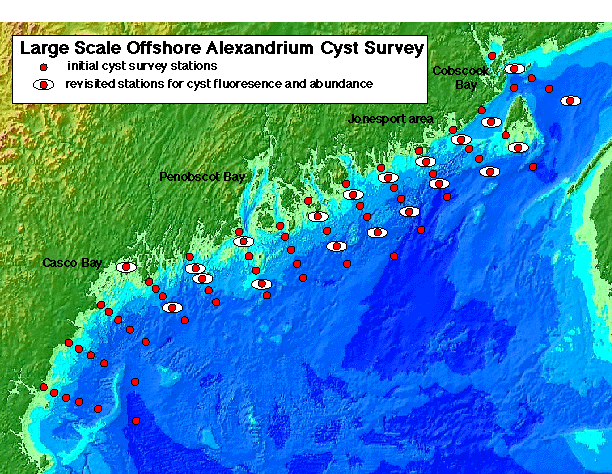
Cyst Mapping. Maps of Alexandrium cyst distribution and abundance are essential. The record of cyst distribution in the GOM, though scattered and outdated, indicates that Alexandrium cysts are widely distributed and abundant (Lewis et al. 1979; Thayer et al. 1983; Anderson and Keafer 1985). To document the present-day distribution, sediment samples will be collected from 75 stations in offshore waters between Portsmouth, NH and the Canadian border during the winter, Year 1 and at an additional 100 inshore stations in Casco Bay, in Cobscook Bay and the Jonesport area of downeast Maine (station maps: http://crusty.er.usgs.gov/ecohab/). The latter areas are the most toxic in eastern Maine. Stations have been determined from sediment deposition maps (see web site) which indicate areas of clay, silt, and thus cyst accumulation, and through the use of historical toxicity patterns. Samples will be collected using either a Craib corer (Craib 1965) or a modified Van Veen grab, both used by the PIs to collect undisturbed sediment samples. The top three cm of sediment will be collected and the material stored and processed according to Anderson et al. (1996). Cysts will be enumerated and abundance plotted and contoured to identify areas of high concentration.
Cyst Dynamics. Cyst maps will be used to select key stations to be visited repeatedly during the bloom season to document excystment dynamics (Fig. 4D). Given the importance of cyst dynamics in ECOHAB-GOM and the difficulty in measuring rates, three approaches will be used to estimate the rate and magnitude of cyst germination. With these estimates and maps of cyst abundance, the magnitude of the input from various areas can be extrapolated. First, 5 replicate cores will be collected at selected offshore and nearshore stations in the WMCC and EMCC during each cruise in Years 1 and 3. Changes in Alexandrium cyst abundance at these stations will provide estimates of cyst germination and formation. Cyst resuspension is a potential problem in shallow areas, but this factor can be distinguished from losses due to germination by monitoring cyst fluorescence (see below). New cysts, with their distinctive contents and pigmentation, can be differentiated (Anderson 1980) and thus the timing and magnitude of cyst deposition can be estimated at these re-visited cyst stations.
Alexandrium cysts synthesize chlorophyll as they prepare to germinate and thus emit red fluorescence (Anderson and Keafer 1985). New cysts also fluoresce but have a distinctive starch content (Anderson, 1980) . Fluorescence measurements can therefore indicate when germination is beginning and when new cysts are being deposited. Germination rates will be estimated by monitoring cyst fluorescence at selected stations in both regions throughout the bloom season following Anderson and Keafer's (1985) methods The percentage of cysts that fluoresce through time will be determined and germination rates calculated as a function of time and location using an empirical relationship between the duration of fluorescence and germination at different temperatures (Anderson, unpub.).
Rate data will also be collected using the germination trap sampler of Ishikawa et al. (1995), to be deployed at two locations in Casco Bay in Years 1 and 3. The trap is an inverted, 10 &m mesh-lined pyramid that covers a known area of the bottom. A hose connection to the surface allows water inside the trap to be removed and screened for Alexandrium vegetative cells. Although cell growth and grazing are potential problems, the frequent sampling planned for Casco Bay will minimize these factors. Examples of this approach are in Ishikawa and Taniguchi (1996).
Estimates of the quantity, timing, and rate of cyst formation will be obtained during the field surveys and process studies based on the relative abundance of planozygotes, determined as in Anderson et al. (1983). Planozygotes (vegetative cells which have undergone sexuality and are about to encyst), are large, and easily identified. Net tows will provide the biomass for these enumerations.
Laboratory Studies. To test for the existence of an annual clock, sediment will be collected from one offshore and one nearshore station in each region and treated according to Anderson and Keafer (1987). The temperature threshold for germination of cysts from each region will be determined using a temperature gradient bar (Watras et al. 1982). A related experiment will expose replicate sediment samples to a stepped temperature increase that simulates seasonal warming in deep and shallow environments. By monitoring cyst fluorescence and germination through time, we can obtain data to supplement field observations. In addition, cultures of cells from the eastern Maine region will be established from cysts and vegetative cells for use in this project and in the future.
{kind=link}
{kind=link}
{kind=link}
{kind=link}