| On "The Four Principles of Adaptation"
As an alternative to the precept that natural selection produces
adaptation, there will be presented here a summary of “The
Four Principles of Adaptation” (Hulburt, 2002). The merit
of the approach in this study is that adaptation is considered to
be the present adjustedness of biota to their environment and to
each other – and, as well, the present lack of adjustedness
of biota to their environment and to each other. For it would be
naïve in the extreme to expect only adaptedness, adjustedness,
everywhere always. Moreover, the precept of natural selection is
ambiguous with respect to adaptation. Thus from Stern (1970) we
have 1): “whatever has been produced by selection is to be
designated as better adapted” has the contrapositive; 2):
whatever is not to be designated as better adapted has
not been produced by selection. 1) and 2) together form
a logical whole. But equally well 1) can be elaborated into a logical
whole without negation as 3): whatever has been produced by selection
is to be designated as better adapted and whatever is to be designated
as better adapted has been produced by selection, so that the selected
is equivalent to the better adapted. In 2) a badly adapted component
of nature is suggested to occur and in 3) no badly adapted component
of nature is allowed. In what follows both 2) and 3) will be of
concern, in the sense that nature does have both well adapted and
badly adapted components in some cases and only well adapted components
in other cases.
But to see the options of these cases a wholly different approach
will be taken. The approach will be that a factual, observational
basis must exist and adaptation must be inferred from it. To this
end the four principles of adaptation of Hulburt (2002) will be
presented and these will be seen to emerge from the factual situations
that accompany the principles.
The First Principle
The first principle is that if two quite different entities occur
under the same condition, then one is adapted and the other is not
adapted to this condition. Thus the warm-blooded vertebrate is adapted
to year-round temperature in temperate regions because it is behaviorally
active year-round (except hibernators), whereas the cold-blooded
vertebrate is not adapted to year-round temperature because it is
not active year-round (plus hibernators).
Put in logically valid form, this is as follows: if x is
in a year-round active animal (a squirrel), then x is in
an adapted animal, one that has adaptedness to year-round temperature
– equivalent to: if x is in a not-adapted animal
(a toad), one which does not have adaptedness to year-round temperature,
then x is in a not year-round active animal. The x
is single from one adapted animal to the other unadapted animal,
each kind integrated by the single property of adaptedness or by
the single property of unadaptedness. The single x integrates
the two opposing kinds (the squirrel kind and the toad kind).
The Second Principle
The second principle is that if one entity occurs under two quite
different conditions, then it is adapted to one condition but is
not adapted to the other condition. Thus the North American forest
is adapted to moist conditions in the east and west but is not adapted
to the non-moist conditions of the semi-arid southwest U.S.A. Put
in logically valid form this is as follows: no matter what y
is chosen, if the forest is adapted to y then y
is a moist condition – equivalent to: for any y if
y is not a moist condition then the forest is not adapted
to y. Briefly: the forest is adapted only to moist conditions
if and only it is not adapted to non-moist conditions. In part the
forest is composed of tall densely packed trees, whether in the
diverse, deciduous forest of Appalachia, or in the spruce forest
of northeastern Canada, or the varied evergreen forest of the Rocky
Mountains and Pacific northwest – all moist regions –
and in part the forest is composed of depauperate, spaced-apart
pinon pine and creosote trees of the semi-arid southwest U.S.A.
It is the view here that the depauperate forest of the southwest
is a mute spokesman of the contrast between the unadaptedness that
it exhibits with the adaptedness that the tall densely packed forest
exhibits. It is the same forest in both places, its contrasting
characteristics being spatially and thus non-contradictorially separated
– just as the same tree species is tall and well-formed away
from the coast but short and gnarled on the windswept coast.
The Third Principle
The third principle is that if one entity is adapted to a second,
then the second is adapted to the first. Thus the white spruce was
adapted to an expanding locale between 12000 and 9000 years ago
in the mid-west of North America, and this locale was adapted to
the spruce. For the sake of logical validity and for driving the
point home, we have: if the white spruce was adapted to its expanding
locale, then this locale was adapted to the white spruce, and if
this locale was adapted to the white spruce, then the white spruce
was adapted to it – equivalent to: spruce was adapted to locale
if and only if locale was adapted to spruce. But of course, the
smallest plant is adapted to the spot where it stands and this spot,
this habitat, is adapted to it. The habitat must be adapted
to the plant, otherwise the plant would not be there. Likewise,
the locale must be adapted to the species, otherwise the species
would not be there. This is the most crucial feature of ecology.
The Fourth Principle
The fourth principle is that if two quite different entities occur
under two quite different conditions, then one is adapted to its
condition and the other is adapted to its condition. For example,
and in logically valid structure: there is diapause or there is
non-diapause in insects; if there is diapause (an insect in an overwintering
larvae stage) then there is winter adaptedness, and if there is
non-diapause (the same insect is in a winged stage) then there is
summer adaptedness; so there is winter adaptedness or summer adaptedness.
We have two mutually exclusive and jointly exhaustive stages –
as we do in leafless and leafy trees, in below ground and above
ground parts of perennial plants, in seeds and plants of annual
plants. And paleontologically and evolutionarily we have a tendency
(an entity) toward many boned toes of paddle limbs of aquatic vertebrates
and this is an adaptation to swimming, and there is a tendency toward
two or three toes instead of five in land vertebrates and this is
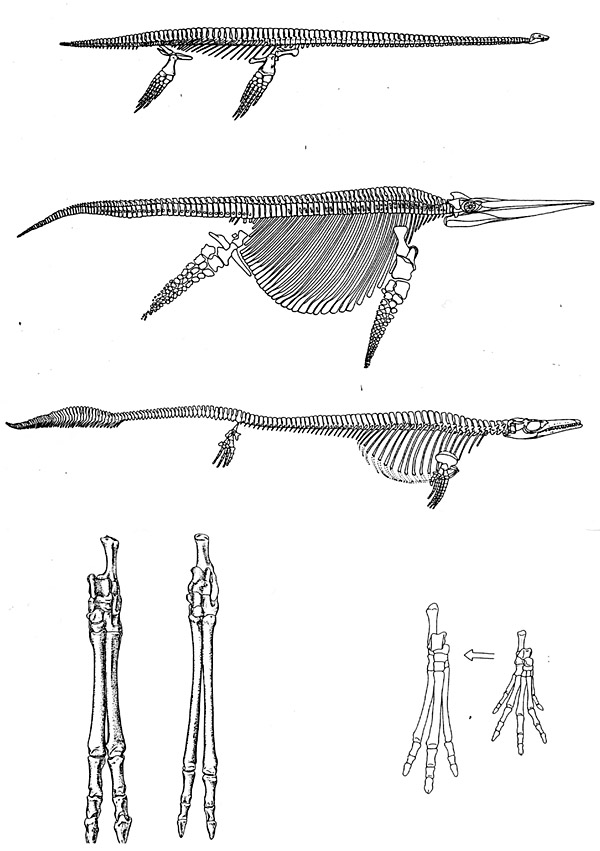
an adaptation to running (Fig. 1). And all of these pairs can be
put in the logically valid structure of the insect example, for
this structure is as much a part of reality as the biota that make
up its content.
So this presentation distinguishes between cases where adaptedness
and non-adaptedness pertain in the first and second principles,
and cases where only adaptedness pertains, in the third and fourth
principles.
The first principle is elaborated in Hulburt (1992), the third principle
is part of the structure of Hulburt (1996, 2000) and the fourth
principle is exemplified by cases in Hulburt (1998).
References
Carroll, R. L., 1988. Vertebrate Paleontology and Evolution. W.
H. Freeman and Company, New York, p. 698.
Hulburt, E. M., 1992. Equivalence and the adaptationist program.
Ecol. Model. 64, 305-329.
Hulburt, E. M., 1996. The symmetry of adaptation in predominantly
asymmetrical contexts. Ecol. Model. 85, 173-185.
Hulburt, E. M., 1998. Theory of adaptation: application of symbolic
logic. Ecol. Model. 107, 35-50.
Hulburt, E. M., 2001. Non-interference and reciprocal adaptation.
Ecol. Model. 2001, 1-13.
Hulburt E. M., 2002. The four principles of adaptation. Ecol.
Model. 156, 61-84.
Stern, J. T., 1970. The meaning of ‘adaptation’ and
its relation to the phenomenon of natural selection. Evol. Bio.
4, 39-66.
Figure 1. Uppermost figure: The plesiosaur
Hydrothecrosaurus, Jurassic, 12 m. long. Next to the top figure:
the icthyosaur Shonisaurus, Upper Triassic, 15 m. long. Middle figure:
the mosasaur Plotosaurus, Upper Cretaceous, 10 m. long. Bottom figure:
left pair, the artiodactyls Proebrotherium and Protoceras, Oligocene;
right pair, the perissodactyls Hyracotherium (left) and Tetraclaenodon
(right), Lower Eocene (from Carroll, 1988, pp. 248, 256, 234, 515,
and 529).
|
